Leaderboard




Popular Content
Showing content with the highest reputation on 10/07/2021 in all areas
-
Hypothetically they could but there are not many examples of parallel evolution taken to the necessary degree. The more similar the end result and the deeper in time the split, the less likely that option becomes. Example would be mara and european hares .. though they evolved to fill a very similar niche on different continents, their differences are as obvious as their similarities. The farther we go back in time for a split from the known pongid/human tree, the more likely that either fossils will exist somewhere or the thing is imaginary ... and the tracks I've cast were real, the plaster impressions were real, and I can tell you that in the setting where I found "my" trackline, a hoax was not possible and nothing exists to misidentify. I think the evidence points to a very late split off the known "tree" or a known split that underwent very rapid physical adaptation due to extreme, extreme climate stress in isolation from the parent population. That isn't something I "know", rather something I compute as best guess based on the evidence available to me. Setting Ketchum aside for a while and focusing on the "theory" of hybridization, the idea has several points of biological merit. First, it has been reported that there are two similar forms, one more apelike with cruder "speech", one more humanlike (in feature) with more articulate "speech". Both are similarly massive but the more ape type is reported as having even more extreme musculature than the other. Second, broad variation in size and color. Third, variations in number of digits, especially toes. Plus other things including native accounts of abducted women who return pregnant with seemingly partially non-human offspring. To me, taken on the whole, it suggests one of two possibilities, either a species in severe decline with a lot of mutations reinforced by massive inbreeding -or- (the option I lean towards) a hybridization event which is still incomplete such that the genes of the 2 parent species are not homogeneously spread across the whole population yet. I find what is happening out there very fascinating. I think there is something quite unique occurring right under our noses that most humans are overlooking. MIB2 points
-
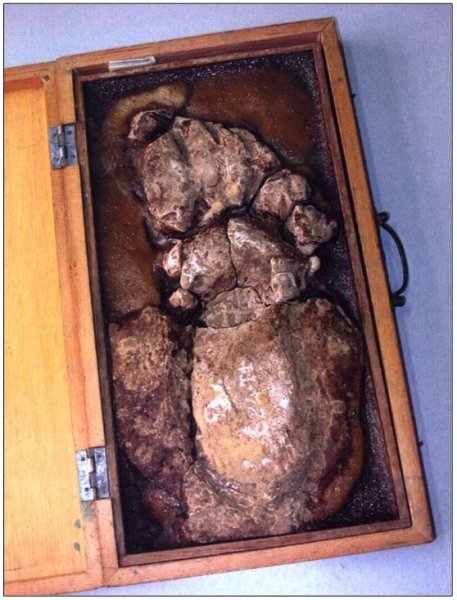
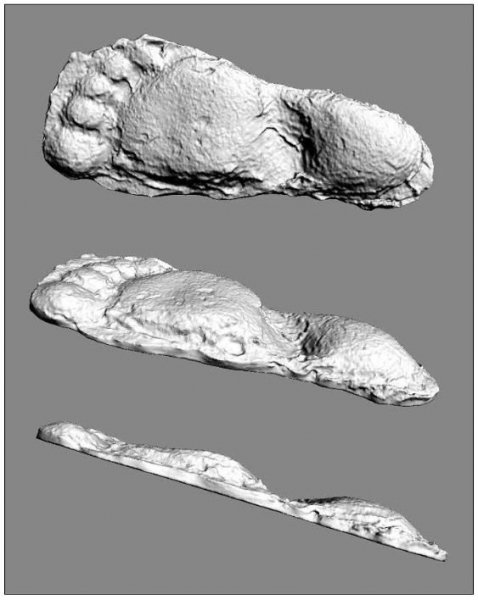
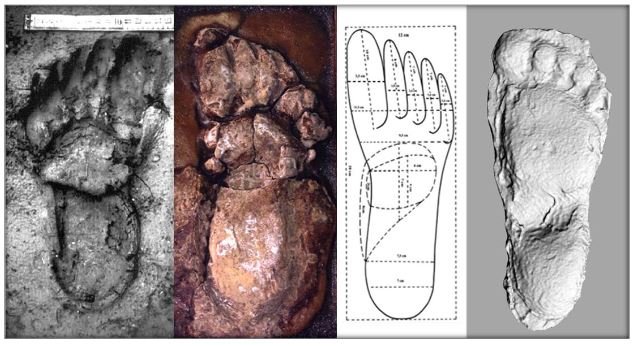
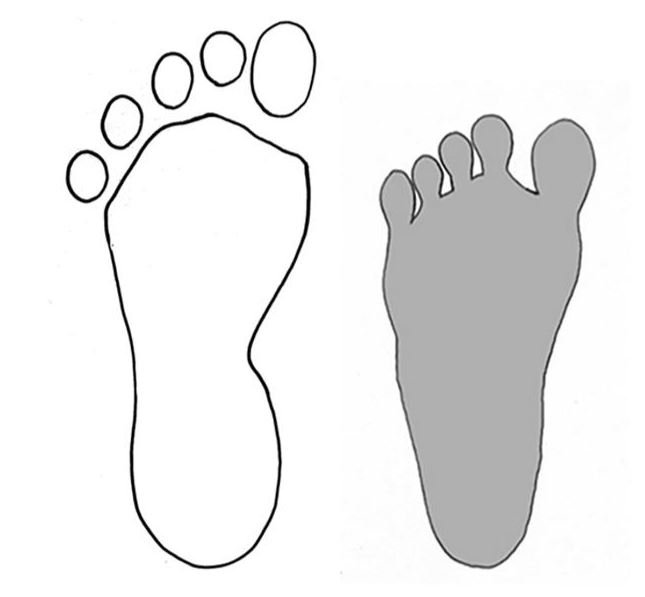
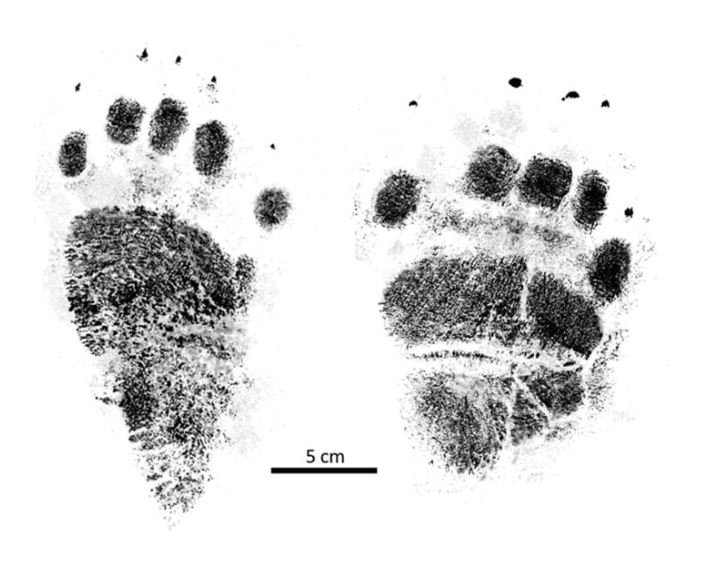
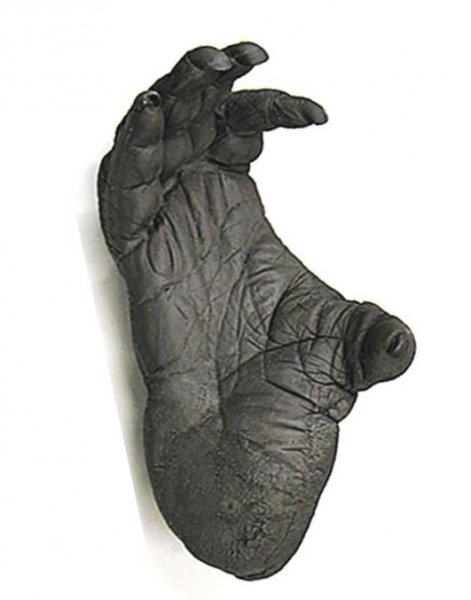
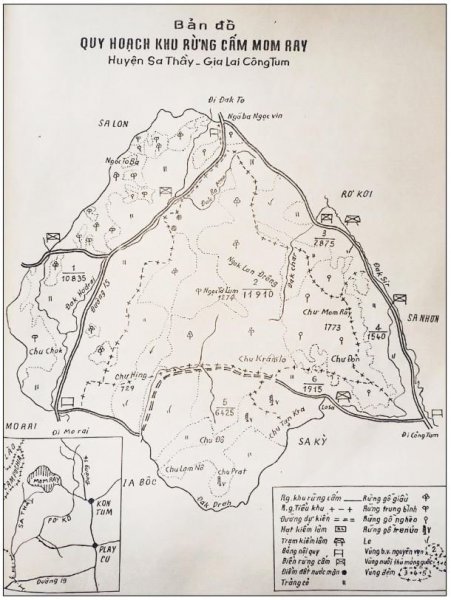
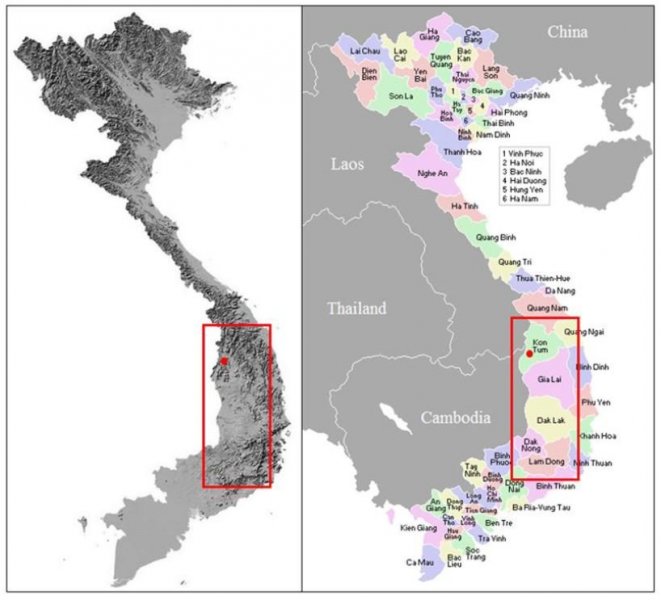
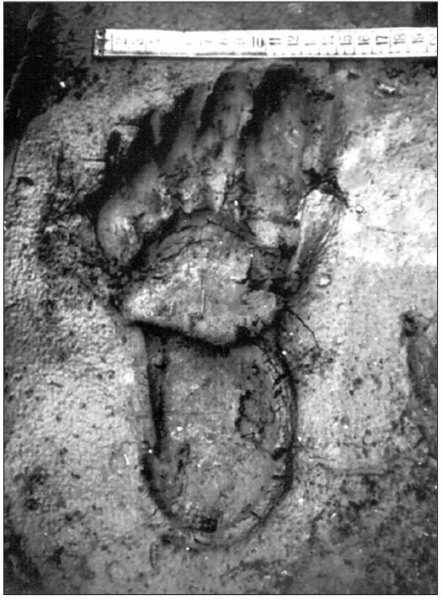
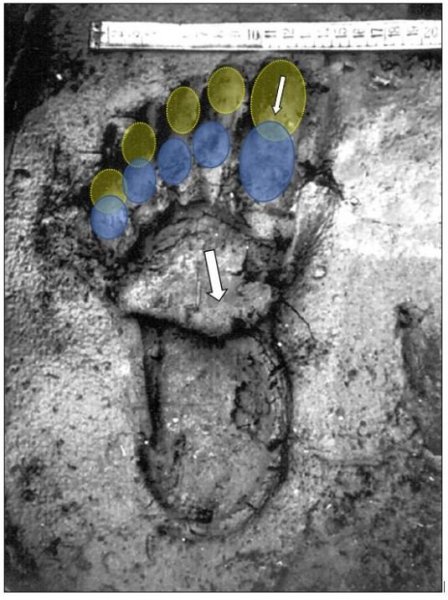
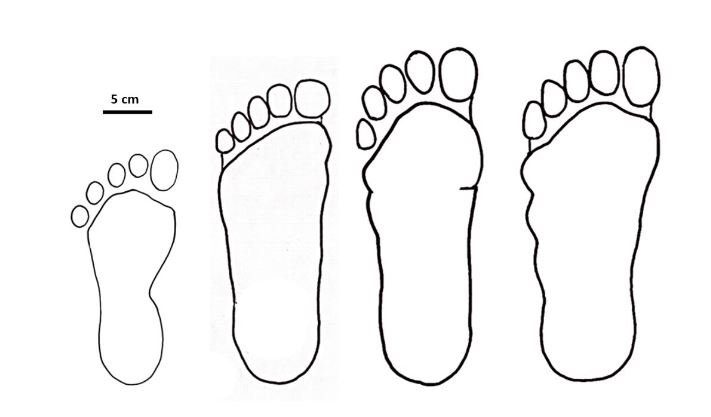
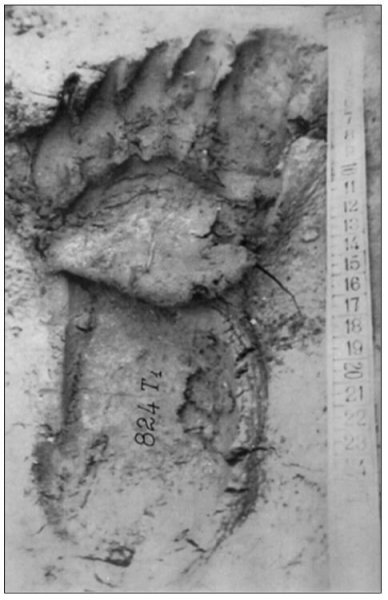
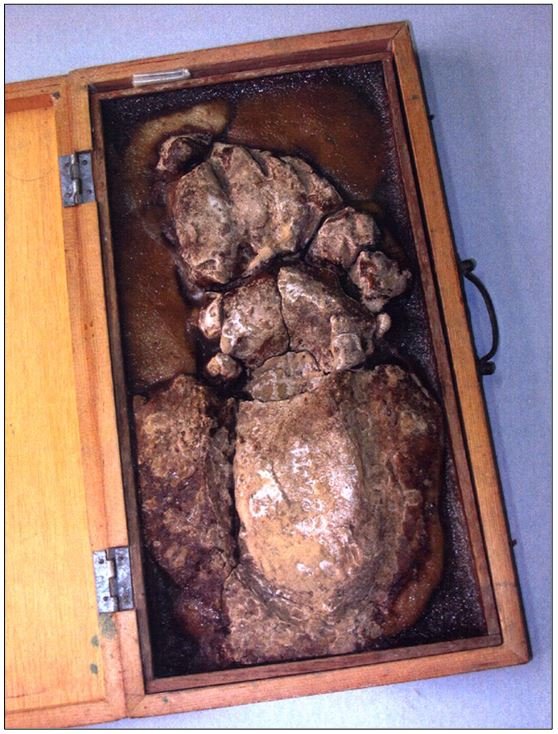
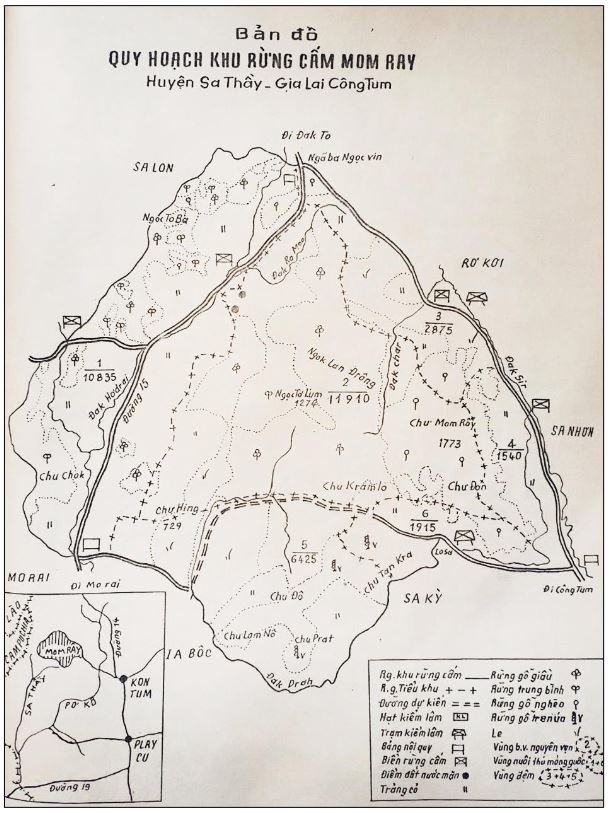
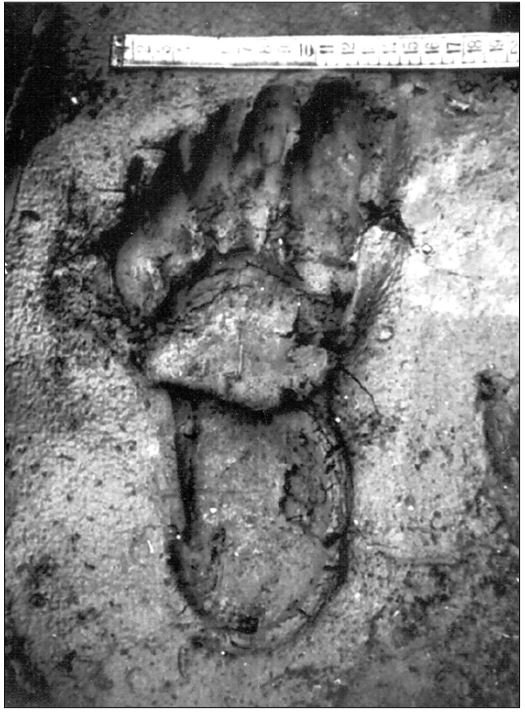
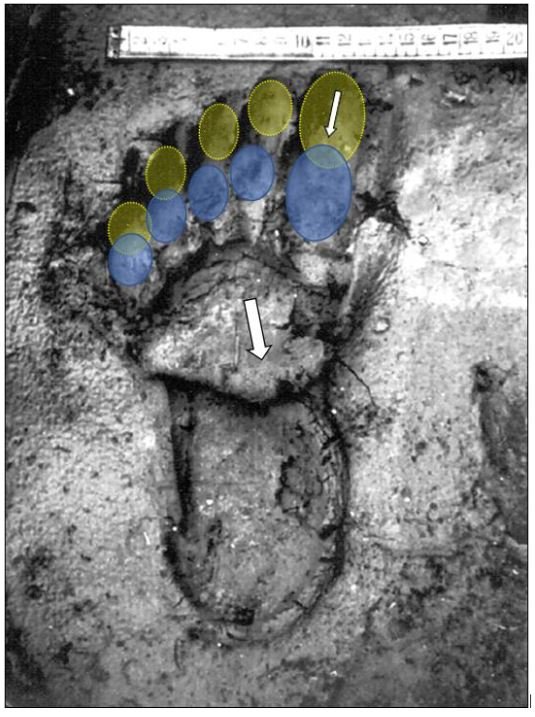
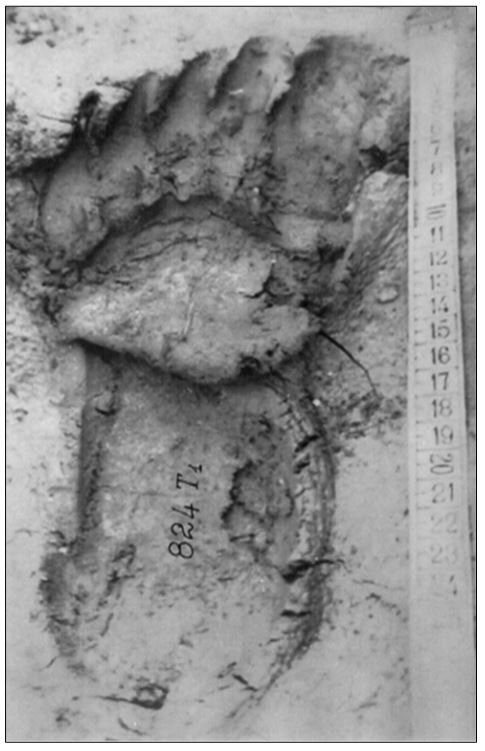
reprinted with permission 10/4/2021 The RELICT HOMINOID INQUIRY 10:6-28 (2021) Research Article RESEARCH ON WILDMEN IN VIETNAM Trần Hồng Việt1*, Trần Hồng Hải1 , Jeff Meldrum2 1 Vietnam Cryptozoic and Rare Animal Research Center, Hanoi University of Education 2 Department of Biological Sciences, Idaho State University, 921 S. 8th Ave, Pocatello, ID 83209 *Correspondence to: Trần Hồng Việt, Email: tranhvietsp@gmail.com © RHI ABSTRACT The possible existence of relict hominoid species in various parts of the world persists as a question of limited overt interest for investigation by most scientists. Investigations in the Central Highlands of Vietnam have documented ethnographic traditions, recent anecdotal descriptions, and trace evidence, i.e. footprints, of possibly two forms of “wildmen” – a great wildman and a small wildman. A documented trackway, providing one fresh hominoid footprint, herein attributed to the great wildman, is larger than a typical local Vietnamese human footprint. Its proportions, combined with indications of a longitudinal arch, are features distinct from the footprints attributed to a sasquatch-like hominoid, as evidenced in both North America and Asia. Given historic impacts on montane habitat presumed to be occupied by the wildmen, their status is likely endangered and timely efforts to identify and understand them are needed. KEY WORDS: Relict Hominoids, Footprints, Indochina, Mainland Southeast Asia, Central Highlands, Người rừng, Sasquatch. INTRODUCTION The existence and nature of relict hominoids, or “wildmen,” is now a topic of interest within the scientific community (Meldrum, 2012a, b). However, research on wildmen attracts the involvement of only a limited number of scientists around the world (Mã Tranh, 2002). In Vietnam, the legends of “forest men” have been recounted for hundreds of years, but it was not until the 1970s that the Vietnamese government showed serious interest in the potential existence of these creatures. In 1974, the Vietnamese government sponsored a research team, which included Prof. Hoang Xuan Chinh (Archaeological Institute), Vo Quy, and Le Vu Khoi (Hanoi University), to go to the Liberated Area of Gialai-Kontum and Dak Lak Provinces, to inquire about the forest men and to gather pertinent information and evidence. Many paleontological and archaeological studies in Vietnam have discovered evidence of primitive hominins (i.e. Homo erectus) in Tham Hai, Tham Khuyen (Lang Son Province), as well as modern humans (Homo sapiens) at Hang Hum (Yen Bai Province), Thung Lang (Ninh Binh Province; Schwartz et al., 1994, 1995; Le Trung Kha, 1974; Ho, PT, 1997). Fossil skeletal remains and tools dating back to the Old Stone Age were found in many places throughout the country (e.g. Nui Do, Nui Nuong, Nui Quan Yen, etc.; Tinh uy, UBND tinh Thanh Hoa, 2000). The latest finding was eleven primitive tools from the Lower Paleolithic Era (about 800 kya) in Roc Tung (An Khe, Gia Lai; Doi et al., 2020 With regard to anthropoids, evidence of Pongo pygmaeus, P. hooijeri, Gigantopithecus blacki, etc. have been found (Schwartz et al., 1995). Lately, two complete subfossil skeletons of orangutan (an adult female and a juvenile), dating back 4,000-5,000 years ago, have been found at Cao Ram (Luong Son, Hoa Binh; Bacon & Vu, 2001). During the two wars in Vietnam (1945- 1975), there were many eyewitness accounts of wildmen-encounters deep in the forest, experienced by American, South Korean and Vietnamese soldiers. Some notable accounts include Captain Frank Hansen’s story of an alleged frozen wildman corpse, eventually exhibited in Minnesota (Huevelmans & Porchnev, 1974; Huevelmans, 2016); Australian journalist Wilfred Burchette’s story about the “forest man” in Dak Mil Province (Central Highlands; see Forth, 2008); Cryptozoologist Loren Coleman’s tale about wildman at Thi Village (Dak Lak; Coleman and Huyghe, 1999; see also Loofs-Wissowa, 1996); Columnist Kregg PJ Jorgenson’s accounts about the Người rừng (Central Highlands; Jorgenson, 2001); and other anecdotal accounts of American GI encounters with “rock apes” (Meldrum, pers. comm.). These persistent reports of encounters with wildmen raised the possibility of the existence a relict hominoid in Vietnam and drew the attention of Vietnamese scientists. FIELD INVESTIGATION Vietnamese scientists had been involved with wildmen investigations since 1974, but it was not until 1977 that the research on wildmen was officially undertaken in the South of Vietnam. In 1982, the research project titled “Study and proposed measures to protect the valuable and rare animals in Sa Thay, Gialai-Kontum” coded 5202-0102b, which is a part of a key national program on environment (5202), was conducted with the main purpose of studying wildmen in the central area of Vietnam (Viet, 1986, 1998). When it was determined that there were indeed some evidences of wildmen in Kon Tum Province, the government resolved (number 65/HDBT, date 7/4/1982) to designate 35,500 ha of forest of Mom Ray Mountain region to protect and study them further. As of 2002, it has been designated the Cum Mom Ray National Park (see Fig. 1, 2). Adjacent to the Chu Mom Ray National Park are protected areas in Cambodia and Loas. The entire area has about 700,000 ha of forests creating a large cross-country preserve, which has particular important to biodiversity conservation in Indochina and Southeast Asia. The region has been identified as the Southern Annamites Montane Rain Forest Ecoregion by the World Wildlife Fund. The intact forests of the ecoregion are wet closed broadleaf evergreen forests receiving up to 200 cm (78 inches) of precipitation. It is otherwise little explored due to its remoteness and hazards (e.g. landmines). The known flora and fauna attest to the region's biological diversity. The tiger (Panthera tigris), Asian elephant (Elephas maximus), douc langur (Pygathrix nemaeus), gibbon (Hylobates gabriellae), wild dog (Cuon alpinus), sun bear (Ursus malayanus), clouded leopard (Pardofelis nebulosa), gaur (Bos gaurus), banteng (Bos javanicus), and Eld's deer (Cervus eldii) are among the better known of the 122 mammal species found here (Wikramanayake, n.d.). Ten field investigations, comprising a total of 432 days, at 21 locations in Gia Lai, Kon Tum, Dak Lak, and Lam Dong Provinces were undertaken (Fig. 2). Eyewitness accounts of the wildmen were gathered from the locals, and a thorough study of environmental habitat, traces, footprints, diet, activities, etc. was made where the locals had reportedly encountered the wildmen. Unfortunately, due to inadequate expedition conditions, the serious deficiency of the equipment, limited time and interruption during the fieldwork, the data collected were not as abundant as expected and no conclusive evidence of the existence of the wildmen was found. There were, however, some noteworthy observations as follows: 1.) Wildmen are suspected to exist in a few scattered, secluded areas in the eastern south-central coast of Vietnam (from Quang Nam to Phu Khanh) and in five provinces in the Western Truong Son Range (Annamite Range). Nevertheless, during and after the wars, the habitat has been shrinking significantly. 2.) There are perhaps two kinds of wildmen co-existing in Vietnam: the “great wildman” and the “small wildman”. Their range may overlap in one area – Dak Lak Province. GREAT WILDMAN Local names: Người rừng (Kinh), Kdghăt (Ê Đê), Jring (Mơ nông), Bông bót, bơ ban mơ nâng (Giarai). The larger form is reported to be about 1.8-2.0 m (5 ft 11 in – 6 ft 7 in) in height. Their body is covered with long red-brown or darkbrown hair, excepting the face, which skin is pink-grey. The hair on their head is unkempt, hanging down to the shoulder or middle of the back, longer than remaining body hair. The hair on their back lies smooth while that on their belly is separated in two directions, parted in the midline of their body, from the neck to the end of the belly (compare Fig. 3). The forearm hair on the outside is long and twirls up toward elbow. The back of their hands and feet are also covered with hair. They have a robust physique and disproportionately thick body. They walk up-right and bipedally at a slow and leisurely pace with a slight stoop, hands hanging down to the knees and fingers rather curved. However, in the face of danger, they can leap and run very fast. They are not afraid of humans, and do not run away unless approached. They are active both diurnally and nocturnally. They have been encountered at 7:00-9:00 am, 1:00 pm, 4:00 pm, 9:00 pm, and some indeterminate times based on discovered footprints, but the locals usually saw them during the daytime (11 out of 18 encounters). They were seen solitarily most of the time. Out of 18 encounters, the locals only saw a three-member group twice, and a male and female couple five times. According to the locals, wildmen are omnivorous, with varied diets including leaves, wild sour berries, rattan sprouts (Calamus bonianus), tiger grass sprouts (Thysanolaena latifolia), pith and fruit of banana (Musa coccinae); also, bird eggs, young birds, small trapped animals, frogs, toads, fish, shrimp, crab, snails, larvae, and insects. There is no evidence of fire use, so food is presumed to be eaten raw. They use simple sounds, such as long or short, monotonous yelling calls, or sometimes wails, to vocalize. There is no evidence of communication using articulate speech. It is noteworthy that no one has reported being attacked by wildmen in eighteen known encounters, which indicates that they are generally not fierce or aggressive towards humans. Footprint Evidence Regarding trace evidence attributed to the great wildman, more than ten human-shaped footprints were discovered on Ngoc Vin pass, Mom Ray Mountain, Sa Thay district, Kontum province, on April 23, 1982. The footprints clearly indicated a bipedal walk. The distance between steps was approximately 70 cm (27.5 in). Regrettably, it was impossible to fully document all these footprints, because they were left on grass, or firm ground of the mountain road covered by a thin layer of dust (Fig. 3). Field investigators were only able to obtain one clear and detailed footprint, likely less than 24 hours old, coded 824 T1 (Fig. 4). It had a depth of 1.5 cm, as it was left on the roadside near an abyss, on fine soft soil moistened by a rain. The footprint indicates that the great wildmen possess a plantigrade, pentadactyl foot. Footprint 824 T1 was 29 x 12 cm (11.4 x 4.7 in) in length x breadth. The footprint presents elongated toe imprints and a broad rounded heel. In general, it resembles human footprints, but is bigger than that of typical modern Vietnamese, including habitually unshod indigenous human populations. The average Vietnamese man’s height is 162 cm (5 ft 3 in) making them the fourth shortest human population in the world (NCDRisC, 2016). That equates to an average foot length of ≈ 24 cm (9.5 in). One of us (DJM) has suggested the possibility of slippage during the latter part of stance may be indicated in the footprint, which would somewhat exaggerate the apparent length of the toes and would account for a prominently extruded pressure ridge proximal to the metatarsal heads (see Fig. 5). However, both THV and THH, who examined and documented the footprint in situ first-hand, assert that no appearance of slippage was evident. As only one distinct footprint was left in mud, we are left with no basis of comparison by which to conclusively distinguish potential footprint artifact from the actual dynamic morphology. Figure 6 depicts a reconstruction of the outline of the foot without the potential distortion resulting from late stance slippage. This contrasts with the outline in Fig. 11, traced directly from the resulting cast of the footprint. The foot length (from heel to the tip of hallux) is 29 ± 0.5 cm (11.4 in), while that of an average Vietnamese only measures ≈ 24 cm(9.5 in). The forefoot is wide, narrows to the hindfoot, with the widest part across splayed toes (between hallux and the outermost toe) measuring 11.5 ± 0.5 cm (4.5 in). The sole pad breadth across the forefoot measures 9 ± 0.5 cm(3.5 in); average Vietnamese ≈ 7 cm (2.8 in); heel breadth is 7.5 ± 0.5 cm (3 in); average Vietnamese ≈ 6 cm ( 2.4 in). The heel has an evenly rounded outline instead of a tapered one. Although footprint 824 T1 is larger than an average Vietnamese male, it is not as big as the footprints attributed to the sasquatch in North America, with an average length of 40 cm, or 15.75 inches (Fig. 7) (Fahrenbach, 1997-1998; Meldrum, 2006, 2007). The potential presence of a sasquatch-like relict hominoid in East Asia is indicated by examples of equivalent footprints, in morphology and dimensions, discovered in various regions, such as those attributed to the Chinese yeren (Meldrum & Zhou, 2012). The footprint 824 T1 shows a non-divergent hallux, which resembles humans. The toes decrease in size from the hallux to the outermost toes, slightly spread in fan-like shape, point forwards but slant rather medially (perhaps due to slippage of the forefoot); the lateral toes are somewhat splayed initially, but become closely appressed to the hallux, as they flexed into the soil. Adjusting for possible distortion, the hallux pad imprint measures ≈4.6 cm long by 3.0 cm wide (1.8 in x 1.2 in); the lateral toepads are approximately 2.5 cm long by 2.0 cm wide (1 in x 0.8 in). The impressions of the toe stems of the first three digits are evident, as in a human-like footprint. The location of the pressure ridge of extruded mud, produced proximal to the imprint of the metatarsal heads indicates the presence of a longitudinal arch, either transient of fixed. That the arch is rather obscured by the extruded pressure ridge, and no additional footprints are available for comparison, establishing whether the arch is fixed or transient, remains uncertain. The medial prominence of the extruded pressure ridge suggests differential plantar pressure exerted through the medial ball and hallux (Fig. 7). This is further indicated by the greater depth under the hallux compared to the lateral digits (Fig. 8). A cast was made of the footprint, which provides additional insights into the differential topography of the footprint (Fig. 9) further indicating the presence of a medial longitudinal arch, differential expression of the metatarsal pressure ridge proximal to the hallux, and differential depth of impression beneath the hallux. The configuration of shape and proportion, as well as the signs of dynamic animation make it unlikely that this footprint is the result of an artificial contrivance. Furthermore, it neither appears to be an ursid hind paw or the registered fore and hind paw of an ursid. Nor the composite of overlapping spoor of other species. No other footprints of a second animal were visible on site, within a 2 m radius. This morphology of 824 T1 stands in contrast to the diagnosis of the footprint form attributed to sasquatch, based on a large sample of footprints attributed to that possible hominoid (Meldrum, 2007). The sasquatch foot is characterized by a flat flexible foot form, with a considerable range of movement in the midtarsal joints (Meldrum, 2004). A pressure ridge is frequently present associated with flexion at the transverse tarsal joint, i.e., calcaneocuboid + talonavicular joints (Fig. 10). The relative position of the pressure ridge, proximal to the metatarsal heads in the case of 824 T1, in contrast to a position proximal to the inferred location of the transverse tarsal joint in the case of the sasquatch (Fig. 11). When the footprint was discovered, alternate hypotheses to account for its attribution were considered: that it was possibly a large human footprint, or it belonged to a bear, or orangutan. It has been pointed out that the feet of the indigenous human populations of the central highlands are notably smaller than 824 T1 (Fig. 12). It seems unlikely that the footprints were made by a barefoot Vietnamese. There are two species of black bear found in southeast Asia, the Asiatic black bear, or moon bear (Ursus thibetanus), and Malaysian sun bear (Helarctos malayanus). These are quadrupeds with distinctive fore and hind paw prints. However, the elongated hind paw and five digital pads lend only a superficial resemblance to a human footprint. Notable distinctions include the tapering pointed heel, usually claw impressions, and reversed appearance of the toe row (the shortest toe is located medially). Members of the IUCN Bear Specialist Group were consulted for an additional opinion concerning the possibility of attributing the footprint to a bear. Asiatic Black Bear Expert Team co-chairs, Dave Garshelis and Mei-hsiu Hwang examined photos of 824 T1 and concluded that it did not convincingly resemble any bear spoor they were familiar with in form or dimensions and offered inked paw prints for comparison (Fig. 13; Meldrum, pers. comm.). The bear hind print in this instance measures 20 cm (7.9 in), compared to footprint 824 T1 at 29 cm (11.4 in). It has also been suggested that the footprint was left by a surviving orangutan, known to have existed on the mainland during the Pleistocene. However, the extant orangutan foot morphology is very different from that of the 824 T1 footprint, because the known extant orangutan is a quadrumanus arboreal hominoid, with very specialized prehensile feet, with a short hallux diverging from and opposing the other extremely elongated toes (Fig. 14). The only subfossil skeletal remains of a mainland orangutan are rather gracile and possess a more extreme intermembral index (165) than the extant species (Bacon & Long 2001). These exceptional limb proportions, with disproportionately elongated forelimbs, are associated with committed arboreal locomotion. Another possibility is that it was a footprint of a surviving Gigantopithecus sp., but unfortunately, the locomotor mode of these hominoids remain unknown in the absence of postcranial skeletal remains. No pedal fossil specimens are available with which to make a comparison. Besides, G. blackii is inferred to have been of enormous dimensions, 200-300 kg (440-660 lb) or more. Extinct hominin species known from the region include Homo erectus and Homo heidelbergensis. The size, shape, and kinematic structure of the footprint 824 T1 indicate that it could reasonably be attributed to the great wildman, confirming descriptions of this potential relict hominoid as a large bipedal terrestrial nonhuman primate, namely a hominin with derived foot morphology, inhabiting the forests of mountainous areas of southeast Asia. SMALL WILDMAN Local names: Người rừng nhỏ (Kinh), Kdjhăt, Jring tan, Arăc tan, Mnuih (Mơ Nông), Hăng háy, Ma lay (Giarai), Dạ tày nông (Xê Đăng), (Ha Lăng). The small wildman form is reportedly 1.2 -1.5 m (3 ft 11 in – 4 ft 11 in) in height on average, tail absent, entirely covered with long grey-brown or grey-black hair, including the backs of the hands and dorsum of the feet. The head hair is said to be about waist-length for females and a bit shorter for males; the old individuals also have white hair. The face is hairless, and the skin is a light yellow-grey or light grey-black color. They walk upright and bipedally, like the great wildmen, but without a stoop, and can also leap and run very fast. The small wildmen are said to be diurnal. Reports of encounters by locals have occurred during the daytime 13 times, but only once at about 9:00 pm and once at 2:00 am out of 15 encounter cases. Small wildmen are said to live in caves and find food (e.g., small crabs, snails and fish) alongside springs and streams, which are eaten raw, as there is no indication of fire use. They socialize in larger groups than the great wildmen, often numbering 3-5 members. The locals described seeing a group of small wildmen twice, a four-member group once, a male and female couple four times, a pair made of mother and juvenile twice, and a solitary individual seven times. Small wildmen display no evidence of articulate speech. Apart from monotonous yelling calls, they are described as communicating by gestures and sound signals during daytime activities. Their opportunistic tool-use compares to other non-human hominoids, such as using rocks for digging and throwing. Interestingly, both kinds of wildmen in Vietnam are described as not fierce, lacking language, fire use or tool manufacture, and are frequently encountered in proximity of human settlement. They have been seen on the hills, in the fields, or on the roads, where people commonly pass nearby. Small wildmen in particular, are not afraid of humans. They do not run away unless approached. Some eyewitnesses also claimed to have seen them entering the locals’ shacks. They are even perceived to be so friendly and human-like that the locals refer to them as “brothers in the forest.” Perhaps with the rapid proliferation of smartphones, photographic evidence of the small wildmen will eventually be forthcoming. CONCLUSION Vietnam harbors much biodiversity, including rare and unique endemic organisms and an unusual mixture of tropical and temperate species (Sterling et al., 2006). The description of the size, shape, and behavior of “wildmen,” based on eyewitness accounts and alleged specimens examined, suggests the existence of two forms of relict hominoid – the great wildman and the small wildman – in some provinces of Tay Nguyen (Vietnam). Evidence indicates the primary habitat of the wildmen to be the forests in mountainous regions of the Central Highlands, stretching from Kom Tun to Lang Don Provinces. The impacts of war and recent settlement, cultivation and development of natural resources, means the habitat has been shrinking considerably, raising concern that both species are being pushed to the verge of extinction. Vietnamese authorities have yet to show appropriate interest, let alone make proper investment of resources and effort in determining the existence of and researching the nature of wildmen, considering their likely endangered status. Literature Cited Bacon A-M and Long V (2001). The first discovery of a complete skeleton of a fossil orang-utan in a cave of the Hoa Binh Province, Vietnam. Journal of Human Evolution. 41:227-41. Coleman L and Huyghe P. The Field guide to Bigfoot, Yeti, and Other Mystery Primates Worldwide. New York: Avon Books, 1999. Doi, Nguyen gia and Doan khai quat hop tac Viet-Nga (2020) Overview on excavation process and research into An Khe Paleolithic site complex Gialai province in 2015-2019. Khao co hoc 3:9-23. Fahrenbach WH (1997-1998) Sasquatch: size, scaling and statistics. Cryptozoology, 13:47-75. Forth G. Images of the Wildman in Southeast Asia: An Anthropological Perspective. New York: Routledge, 2008. Ho PT (1997) Nguon goc loai nguoi, Nxb Giáo Dục, 75 tr. Heuvelmans B and B Porchnev. L’homme de Neanderthal est toujours vivant, Paris: Plon, 1974. Huevelmans B. The Strange Saga of the Minnesota Iceman, translated by Paul LeBlond, San Antonio: Anamolist Books, 2016. Jorgenson KPJ, Very Crazy G.I., New York: Presidio Press, 2001. Kha LT (1974) Tim hieu van de Neanderthal. Khao co hoc so 16 / 1974: 31-36. Loofs-Wissowa H, “Seeing is believing or is it? How scientific is ‘Wildman’ research?” ANU Reporter 27(12):4 (17 July 1996). Mã T (2002) (translated by Nguyen Duy Chiem). Bi an ve Nguoi rung, Hanoi: Hanoi Publishing House, 335p. Machusin, GN (1986) Nguon goc loai ngưoi (Phạm Thai Xuyen dịch), Nxb KHKT Hanoi, 240p. Meldrum DJ (2004) Midfoot Flexibility, Fossil Footprints, and Sasquatch Steps: New Perspectives on the Evolution of Bipedalism. Journal of Scientific Exploration 18:65-79. Meldrum J. Sasquatch: Legend Meets Science, New York: Tom Doherty Associates, 2006. Meldrum, DJ (2007) Ichnotaxonomy of giant hominoid tracks in North America. In: SG Lucas, JA Spielman and MG Lockley (eds) Cenozoic Vertebrate Tracks and Traces. New Mexico Museum of Natural History and Science Bulletin 42:225-331. Meldrum DJ (2012a) Adaptive radiations, bushy trees, and relict hominoids. The Relict Hominoid Inquiry 1:51-56. Meldrum DJ (2012b) Are other hominins (hominoids) alive today? The Relict Hominoid Inquiry 1:67-71. Meldrum DJ and G Zhou (2012) Footprint evidence of the Chinese yeren. The Relict Hominoid Inquiry 1:57-66. NCD Risk Factor Collaboration (NCD-RisC). “A century of trends in adult human height.” eLife vol. 5 e13410. 26 Jul. 2016. Schwartz JH, VT Long, NL Cuong, LT Kha, I Tattersall (1994) A diverse Hominoid fauna from the late middle Pleistocene breccia cave of Tham Khuyen, Socialist Republic of Vietnam Anthropological Papers of the American Museum of Natural History, Number 73, 11p. Schwartz JH, VT Long, NL Cuong, LT Kha, I Tattersall (1995) A review of the Pleistocene Hominoid fauna of the Socialist Republic of Vietnam (excluding Hylobatidae). Anthropological Papers of the American Museum of Natural History, Number 76, 23p. Sterling E, M Hurley and M Le (2006). Vietnam: A Natural History. New Haven: Yale University Press. Tinh uy, UBND, HDND tinh Thanh Hoa, 2000. Thanh Hoa thoi tien su, Dia chi Thanh Hoa, tap I, Nxb Van Hoa Thong Tin: p. 519-524. Viet TH (1986) Thu hoang dai vung Sa Thay va y nghĩa kinh te cua chung. Truong DHTH Hanoi, 225p. Viet TH (1998) Báo cáo nghiệm thu de tai Nguoi rung, DHQG, DHSP HN: 25p. Wikramanayake E, P Rundel and R Boonratana (n.d.) Southeastern Asia: Vietnam into Laos and Cambodia. https://www.worldwildlife.org/ecoregions/im0152 (retrieved 4/8/2021. WWF Dong Duong (2000) Gioi thieu mot so loai thu o Dong Duong va Thai Lan, 255p. Figure 1. Map of the forbidden forest of Mom Ray (Sa Thay, Gia Lai, Kon Tum Province). Figure 2. Kon Tum, Gia Lai, Dak Lak, Dak Nong and Lam Dong Provinces (Central Highlands of Vietnam). Chu Mom Ray National Park indicated by the red dot. Figure 3. Descriptions of the great wildman resemble this rendition of “Pongoid Man,” by Alika Lindbergh, based on Heuvelmans’ reconstruction of the Minnesota Iceman. Figure 4. The site at Ngoc Vin pass, Mom Ray Mountain, Sa Thay district, Kontum Province, where the trackway trace attributed to the great wildman was left on a mountain road. X marks the location of the single footprint 824 T1. Figure 4. Footprint 824 T1, attributed to the great wildman. Figure 5. Footprint 824 T1. Alternate interpretation of the footprint, allowing for distortion dues to slippage during the late stance phase (DJM). Yellow indicates the initial position of the toe pads in early stance; blue the final position of toe pads after inferred slippage (smaller arrow). Direction of extrusion feature proximal to metatarsal heads (larger arrow) Figure 6. A reconstructed outline of footprint 824 T1 (left), compared to three examples of outlined footprints attributed to sasquatch in North America (right). Figure 7. Footprint 824 T1. Oblique proximodistal view. Figure 8. Footprint 824 T1. Details of the forefoot viewed proximodistally. Note differential depth of the hallux imprint. Figure 9. Cast of footprint 824 T1. Figure 10. Multiple views of a 3D scan of a footprint cast attributed to sasquatch, made by Bob Titmus at the Patterson-Gimlin film site in northern California, 1967, illustrating a midtarsal pressure ridge. Figure 11. (Left to Right) Footprint 824 T1; Cast of 824 T1; Diagram of measurements taken from the cast of 824 T1; Cast of a footprint attributed to sasquatch from the Patterson-Gimlin film site, California. Figure 12. Left. A reconstructed outline of the footprint 824 T1, removing potential distortion caused by slippage (29 cm). Right. Habitually unshod human footprint (24 cm). Figure 13. (Left to Right) Inked footprints of the left hind paw and left fore paw of an Asiatic black bear, Ursus thibetanus (courtesy of Garshelis and Mei-hsiu). Figure 14. Life mold of an orangutan right foot (credit: Bone Clones).



 1 point
1 point -
(Copied / pasted) THE EARLIEST KNOWN MATING BETWEEN DIFFERENT HUMAN SPECIES has been defined by ancient DNA analysis. This interspecies sex happened about 700,000 years ago, long before we (Homo sapiens) were around. Ancestors of both Neanderthals and Denisovans interbred with their Eurasian predecessors - a well-established "superarchaic" population that had separated from other humans 2 million years ago. The time frame fits with the mystery sex partner being Homo erectus, who has to be regarded as the most successful human species of all time, lasting for 1.8 million years compared to our ~300,000 years to date. It's AMAZING what else the ancient DNA analysis can tell us: The superarchaic population was large (between 20 and 50 thousand individuals). When Neanderthals interbred with the superarchaics, their populations had been separate for 1.2 million years. Later, when this mystery population exchanged genes with Denisovans, the two populations had been separated even longer. This made them the most distantly related humans ever to interbreed. The DNA analysis also indicated that ancestors to Neanderthals and Denisovans endured at least one significant population bottleneck. It's mind blowing what DNA analysis is uncovering - stuff we would NEVER know otherwise! Here's the article that also has a link to the Rogers et al. (2020) research paper: https://www.businessinsider.com/ancient-humans-interbreed-with-mysterious-population-2020-2?fbclid=IwAR2EFwmtt3fzsvYamMHgfSm3VqQG00WqZmocx7gOcC_m0XQiU8pkrinu2VI1 point
-
I thought the report was rather weak with the line regarding hypertrichosis and no mention of Zana's other features. I'll repeat my reply to Twist: I wasn't sure. That's why I asked. A well thought out opinion on all wild man reports being people suffering from hypertrichosis isn't likely to be published by a "scientist", but it's going to be a pretty standard escape clause in every case like this that comes along. Let's face it: if a sasquatch (almas, yeti, et al) report cannot possibly be a mistaken bear sighting, it will be a human with hypertrichosis until proven otherwise, even if said hirstute bipedal creature can leap tall buildings, swim flood stage frozen rivers, sleep outside in sub-zero temps without bedding, and carry great weights with one hand. Thank you for that information, and I apologize for coming on strong. That's simply how I am. I guess I'm like Zana, but with just a slight case of hypertrichosis. I swim underwater in frozen lakes, am (somewhat) bullet proof, and am almost as strong as Zana, but have great suspicion of people from the International Skeptics Forum, formerly known as JREF. I will review the forum archives to review your history. Your post, without much comment, was suspicious to me. Yes, the Zana story is an incredible one, even if she was herself or was a descendant of African slaves. If she was simply a feral human "with issues" (as Twist tried to pass), the concept still poses problems. And since East African slavery in that part of the world goes back at least a thousand years, even this DNA review leaves open the possibility of a repressed human population going feral by necessity, even if they're not hybrid with ancient hominid species. In fact, a population of humans hiding from other humans for so many generations as to begin exhibiting the kinds of differences that Zana showed is even more remarkable (and believable) than a surviving ancient hominid or a hybrid sapien/hominid. And, again, it intensifies the ethical side of this entire phenomenon.1 point





This leaderboard is set to New York/GMT-04:00